La peau, les poils, les crins sont colorés grâce à des pigments. Ils sont 2 :
L’eumélanine de couleur noir/brun foncé
La phéomélanine de couleur jaune /rouge
Ces pigments sont synthétisés par des cellules de la peau appelées mélanocytes. La production de ces 2 pigments est maîtrisée par 2 gènes. La synthèse d’eumélanine et de la phéomélanine dépend de l'activation/inactivation de plusieurs molécules :
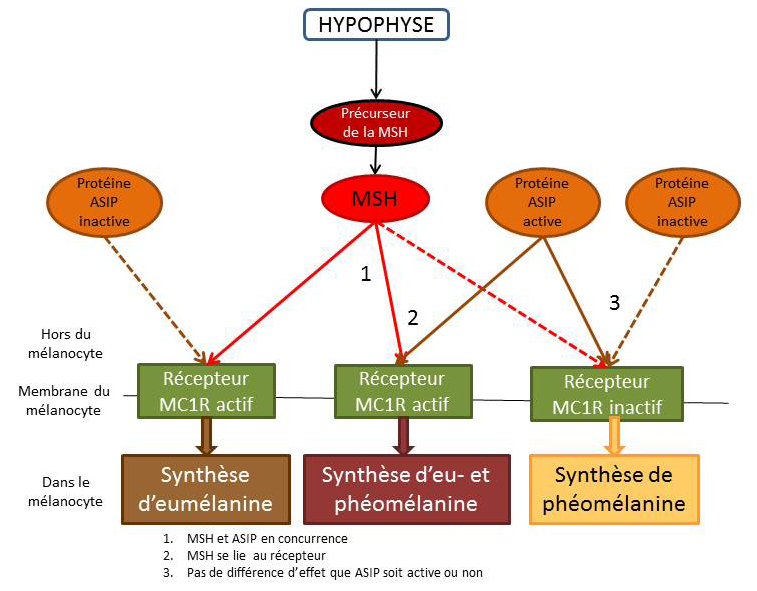
Le récepteur pour MC1R
La protéine ASIP (Agouti Signaling Protein)
Lorsqu'elle est active, elle joue un rôle en se fixant sur le récepteur MC1R (Melanocortin 1 Receptor). Cela entraîne la synthèse de la phéomélanine.
Lorsqu'elle est inactive, le récepteur MC1R peut être activé par un signal hormonal: MSH (Melanocortin Stimulating Hormon). Cela entraîne la synthèse d'eumélanine en plus de celle de la phéomélanine.
En l’absence de signal de l'ASIP et de MSH, de la phéomélanine est synthétisée. Les combinaisons fonctionnelles de la protéine ASIP et du récepteur MC1R vont entraîner des modifications drastiques de la couleur de la robe. Ils vont agir comme des interrupteurs ou des modulateurs de production de chacun des deux pigments.
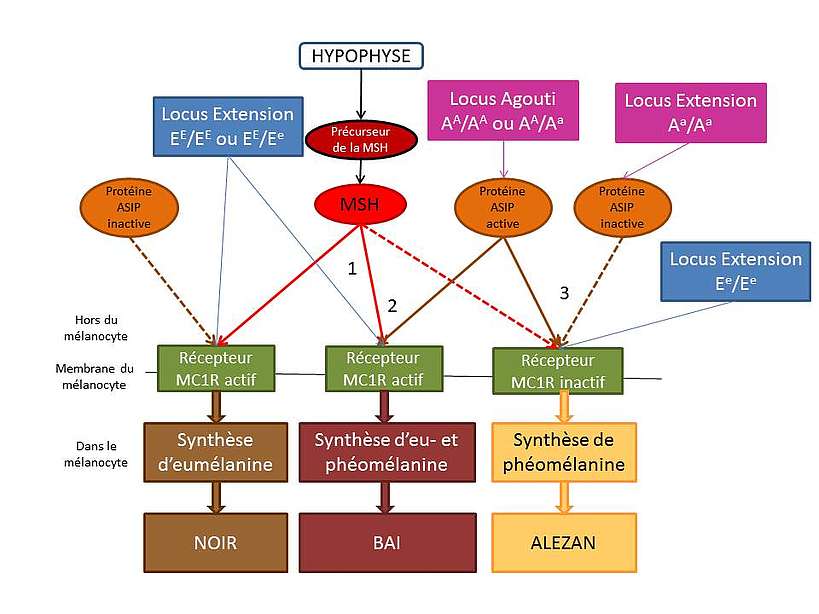
Les couleurs de base sont le bai, l’alezan et le noir. Nous allons considérer que ces trois couleurs résultent essentiellement de l’action de deux gènes : Extension et Agouti.
Gènes intervenant dans la coloration des robes de base
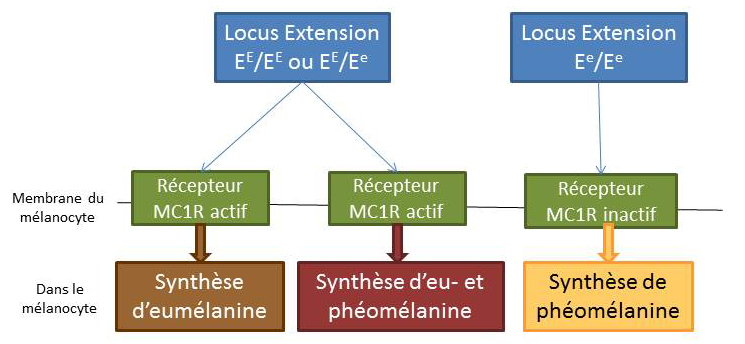
Le gène Extension
Le gène extension code pour l’activation et l’inactivation du récepteur MC1R.
L’allèle dominant sauvage EE
Il code pour un récepteur MC1R actif. Son expression se caractérise de la production d’eumélanine et de phénomélanine.
L’allèle récessif Ee
Il est présent en 2 exemplaires et code pour un récepteur MC1R inactif. Son expression se caractérise par une absence de production d’eumélanine au mélanocyte, seule la phénomélanine est synthétisée.
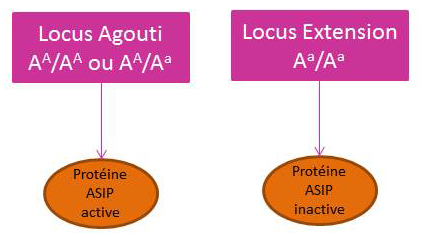
Le gène Agouti
Le gène Agouti code pour l’activation et l’inactivation de la protéine ASIP.
L’allèle dominant sauvage AA
Il code pour une protéine ASIP active. Son expression se caractérise par des extrémités (crins et membres) foncées, c’est-à-dire la restriction de la production d’eumélanine en ces points.
L’allèle récessif Aa
Il est présent en 2 exemplaires et code pour une protéine ASIP inactive. Par conséquent le récepteur MC1R n’est plus régulé et envoie des signaux forts. Son expression se caractérise par une coloration uniforme de la robe : production élevée d’eumélanine au mélanocyte.
Gène Extension
Gène Agouti
Allèle dominant sauvage EE
=> Robe foncée
Allèle dominant sauvage AA
=> Extrémités et crins foncés
Allèle Ee récessif
=> Robe claire
Allèle Aa récessif
=> Robe unie
Génétique des robes de base
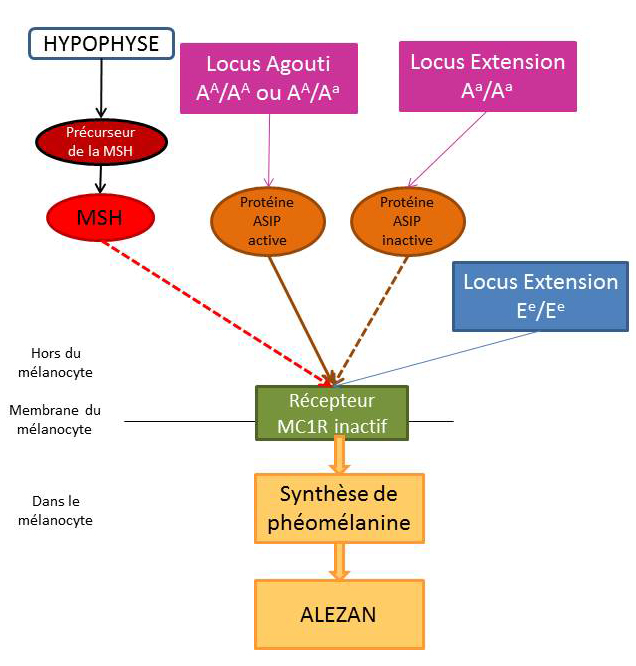
La robe « Alezan »
Pour les chevaux alezans, seule la phéomélanine est produite, résultat d’un récepteur MC1R non actif. Par conséquent, ces chevaux doivent posséder obligatoirement les deux allèles mutés pour le gène Extension : ils sont donc homozygotes récessifs (Ee/Ee).
Le récepteur MC1R étant inactif, la protéine ASIP peut être active ou inactive, cela n’a pas de conséquence. Ainsi, quelle que soit la combinaison des allèles du gène Agouti (AA/Aa , Aa/Aa ou AA/ AA), l’individu homozygote Ee/Ee sera de robe alezane.
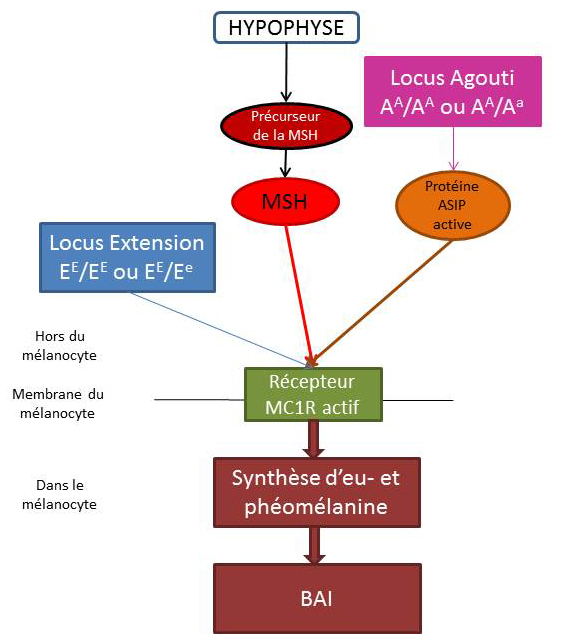
La robe « Bai »
Pour les chevaux bais, la phéomélanine et l’eumélanine sont produites. Les récepteurs MC1R sont donc actif pour pouvoir produire les 2 pigements. Les chevaux doivent posséder obligatoirement un allèle sauvage pour le gène Extension : ils sont donc homozygotes sauvage (EE/EE) ouhétérozygotes (EE/Ee).
Sur les chevaux bais, il y a une variation de couleur au niveau des crins et des extrémités. La protéine ASIP est donc active. Ils doivent donc posséder obligatoirement un allèle sauvage pour le gène Agouti : ils sont donc homozygotes sauvage (AA/AA) ou hétérozygotes (AA/Aa).
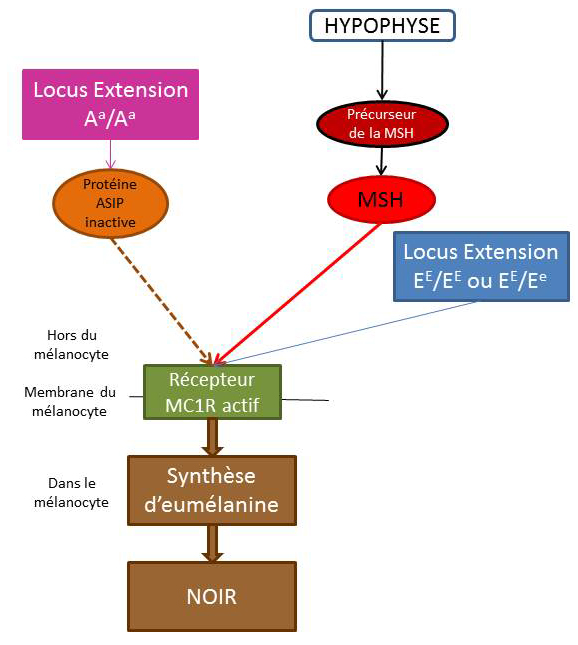
La robe « Noir »
Sur les chevaux noirs, il n’y a aucune variation de couleur au niveau des crins et des extrémités. La protéine ASIP est donc inactive. Ils doivent donc posséder obligatoirement les deux allèles mutés pour le gène Agouti : ils sont donc homozygotes récessifs (Aa/Aa).
Chez ses chevaux, le récepteurs MC1R est actif pour pouvoir produire l’eumélanine. Ils doivent posséder obligatoirement un allèle sauvage pour le gène Extension : ils sont donc homozygotes sauvage (EE/EE) ou hétérozygotes (EE/Ee).
Un test moléculaire a été mis au point pour détecter les individus porteurs de «l’allèle noir» en simple (AA/Aa) ou double exemplaire (Aa/Aa). On peut ainsi distinguer les individus véritablement noirs, de ceux bai très foncés ou alezans brûlés.
Grille de croisement
Si on ne connait pas le génotype des reproducteurs
On a alors une grille de croisement qui prend en compte tous les cas de figures.
Père
Noir
Bai
Alezan
Mère
Noir
93,75% Noir 6,25% Alezan
23,44% Noir 70,31% Bai 6,25% Alezan
37,5% Noir 37,5% Bai 25% Alezan
Bai
23,44% Noir 70,31% Bai 6,25% Alezan
5,86% Noir 87,89% Bai 6,25% Alezan
9,38% Noir 65,63% Bai 25% Alezan
Alezan
37,5% Noir 37,5% Bai 25% Alezan
9,38% Noir 65,63% Bai 25% Alezan
100% Alezan
Ceci confirme la règle d’incompatibilité de filiation, basées sur la couleur : deux parents alezans ne peuvent engendrer qu’un produit alezan.
Si on veut avoir une grille de croisement plus précise, il faut connaître le génotype d’un ou des 2 reproducteurs. Les tests sont très simples, bon marché. Ils se font à partir d’un échantillon de crins ou de sang.
Si on connait le génotype d’au moins 1 des 2 reproducteurs
Père
Bai
Noir
Alezan
AA/AA
EE/EE
AA/Aa
EE/EE
AA/AA
EE/Ee
AA/Aa
EE/Ee
Aa/Aa
EE/EE
Aa/Aa
EE/Ee
AA/AA
Ee/Ee
AA/Aa
Ee/Ee
Aa/Aa
Ee/Ee
Mère
Bai
AA/AA
EE/EE
100% bai
AA/Aa
EE/EE
100% bai
25% noir
75% bai
100% bai
25% noir
75% bai
50% noir
50% bai
100% bai
25% noir
75% bai
50% noir
50% bai
AA/AA
EE/Ee
100% bai
25% alezan
75% bai
100% bai
25% alezan
75% bai
50% bai
50% alezan
AA/Aa
EE/Ee
100% bai
25% noir
75% bai
25% alezan
75% bai
18,75% noir
56,25% bai
25% alezan
50% noir
50% bai
37,5% noir
37,5% bai
25% alezan
50% alezan
50% bai
12,5% noir
37,5% bai
50% alezan
25% noir
25% bai
50% alezan
Noir
Aa/Aa
EE/EE
100% bai
50% noir
50% bai
100% bai
50% noir
50% bai
100% noir
100% bai
50% noir
50% bai
100% noir
Aa/Aa
EE/Ee
100% bai
50% noir
50% bai
25% alezan
75% bai
37,5% noir
37,5% bai
25% alezan
100% noir
25% alezan
75% noir
50% alezan
50% bai
25% noir
25% bai
50% alezan
50% noir
50% bai
Alezan
AA/AA
Ee/Ee
100% bai
50% noir
50% alezan
100% bai
50% bai
50% alezan
100% alezan
AA/Aa
Ee/Ee
100% bai
25% noir
75% bai
50% bai
50% alezan
12,5% noir
37,5% bai
50% alezan
50% bai
50% noir
25% noir
25% bai
50% Alezan
100% alezan
Aa/Aa
Ee/Ee
100% bai
50% bai
50% noir
50% bai
50% alezan
25% noir
25% bai
50% Alezan
100% noir
50% bai
50% noir
100% alezan
En savoir plus sur nos auteurs
Sophie DANVY Ingénieur de développement IFCE
Clothilde DUBOIS Formatrice IFCE
Gérard GUERIN Inra
Margot SABBAGH Ingénieure de projets & développement « Génétique des équidés » IFCE